Persistent Effects
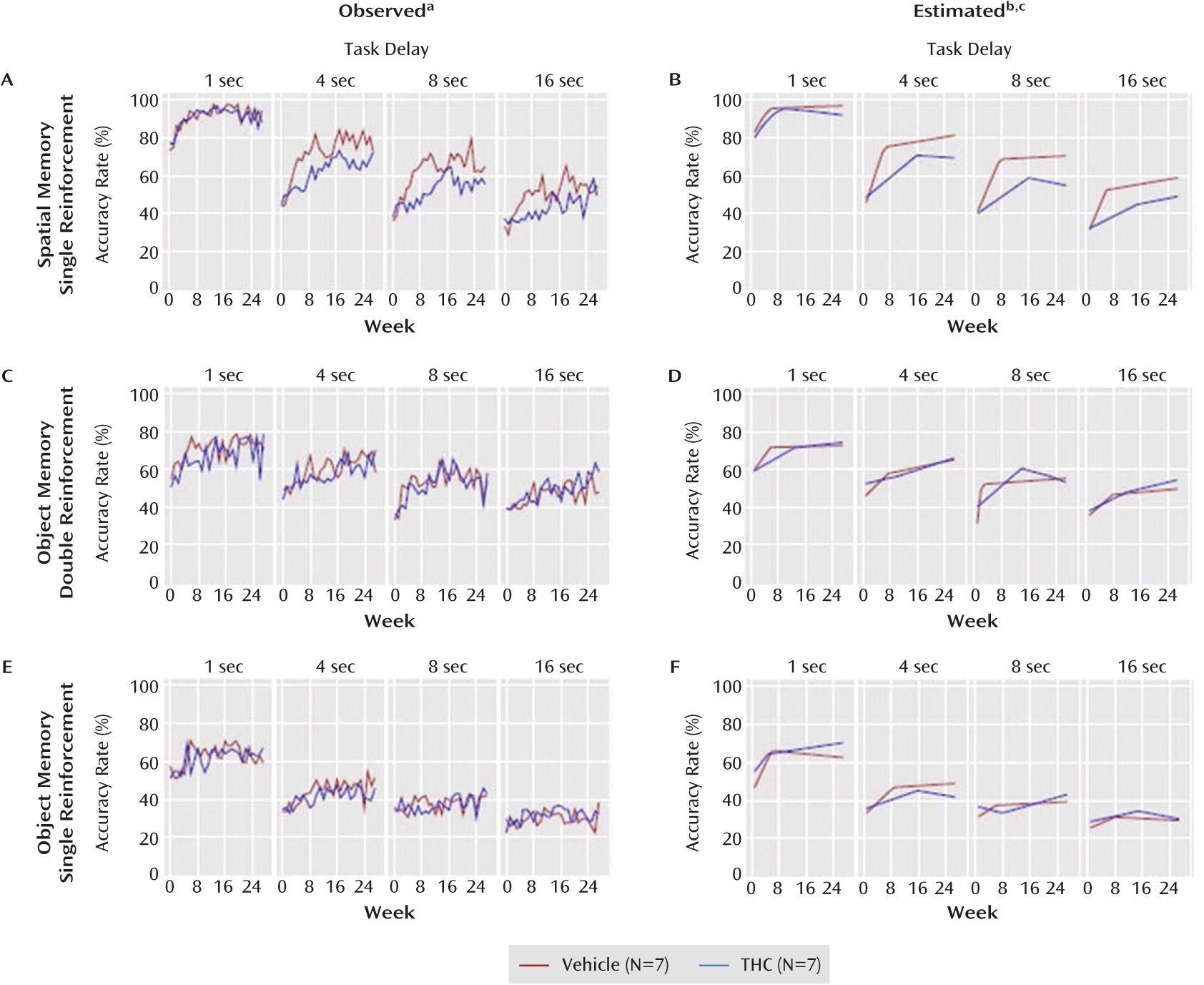
Figure 3 parts A and B display observed and estimated accuracy rates over time for the spatial working memory trials. Accuracy rates increased significantly for both groups on all delays during phase 1 (t≥4.83, df=13, p≤0.001 in all cases). However, the improvement during phase 1 was significantly slower for the THC group on the 4-, 8-, and 16-second delays (t≤–4.16, df=13, p≤0.001 in all cases). Additionally, the length of phase 1 was significantly longer for the THC group by 3.2, 9.5, and 8.4 weeks for the 1-second (t=2.60, df=13, p=0.03), 4-second (t=5.10, df=13, p<0.001), and 8-second (t=4.34, df=13, p=0.001) delays, respectively. For the 16-second delay, the difference was 9.0 weeks (t=1.59, df=13, p=0.14). During phase 2 the only significant slope effects were on the 1-second delay for the THC group, which was negative (t=–2.37, df=13, p=0.04), and on the 4-second delay for the vehicle group, which was positive (t=2.67, df=13, p=0.02). The estimated difference between the THC and vehicle groups at the change point (
Figure 3 part B) was negative for all delays but not significant (p>0.14 in all cases), and the phase 2 slopes (i.e., the rate of change in accuracy between the change point and week 27) did not significantly differ for the 4-, 8-, or 16-second delay (p>0.19 in each case). However, for the 1-second delay the difference was significant (t=−2.63, df=13, p=0.03).
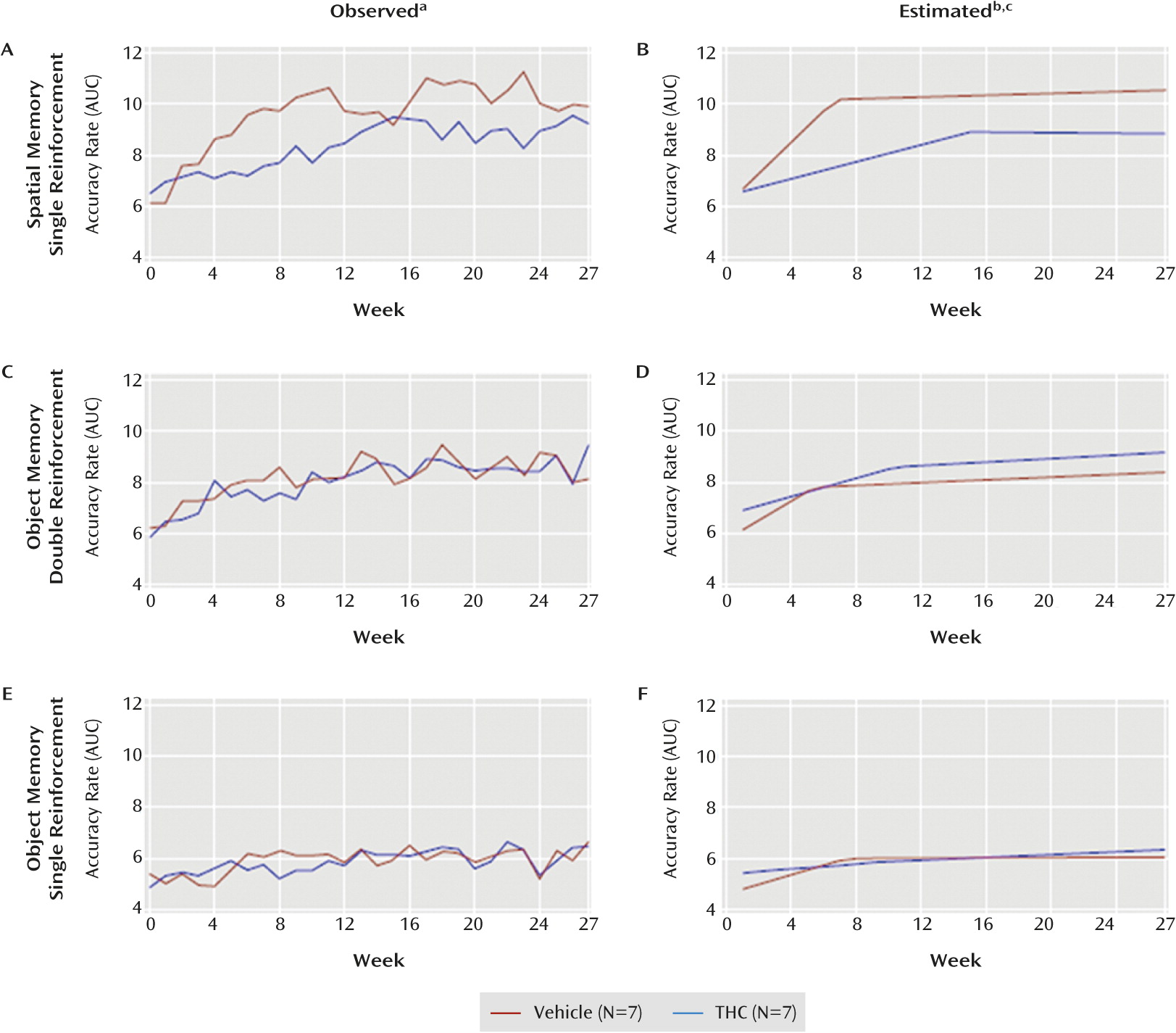
Figure 4 parts A and B display observed and estimated AUCs for the spatial task. Compared with vehicle, THC 1) significantly slowed the rate of improvement during phase 1 (t=–2.76, df=12, p=0.02), 2) significantly extended (by 8.4 weeks) the length of phase 1 (t=2.59, df=12, p=0.03), 3) significantly lowered accuracy at the change point between phases (t=–2.60, df=12, p=0.03), but 4) did not significantly affect phase 2 slopes (p=0.71).
In order to compare performance across delays, we fit a model that included all four delays. The difference between change points was significantly smaller for the 1-second delay than for the 4-second (t=2.53, df=55, p=0.02) and 8-second (t=2.43, df=55, p=0.02) delays. For technical reasons, the 16-second delays could not be compared (see the supplemental results in the online data supplement).
Figure 3 parts C and D display observed and estimated accuracy rates for the double-reinforcement object working memory trials. Accuracy rates increased significantly (t≥2.27, df=13, p≤0.04 in all cases) for both groups during phase 1 on all delays except the 1-second delay for the vehicle group (t=2.14, df=13, p=0.06) and the 4-second delay for the THC group (p=0.24). The only significant difference between groups was on the 8-second delay. Compared with vehicle, THC 1) significantly slowed the rate of improvement during phase 1 (t=–3.09, df=13, p=0.009), 2) significantly extended the length of phase 1 (t=6.50, df=13, p<0.001), but 3) did not significantly affect accuracy at the change point between phases (p=0.18), and 4) did not significantly affect phase 2 slopes (p=0.13).
Figure 4 parts C and D display observed and estimated areas under the curve for the double-reinforcement object memory trials. These data revealed no significant differences between groups. The model with all four delays showed that the difference in change points between groups did not depend on delay. These results indicate that THC had no consistent effects on performance of the double-reinforcement object working memory trials.
The observed and estimated accuracy rates (
Figure 3 parts E and F) and AUC (
Figure 4 parts E and F) for the single-reinforcement object working memory trials also indicate that THC had no consistent effects on performance in these trials (see supplemental results in the online
data supplement).
Performance on the other six performance measures did not differ between the THC and vehicle groups, and there were no apparent trends in p values (see Supplemental Tables 1–6 in the online data supplement).
Acute Effects
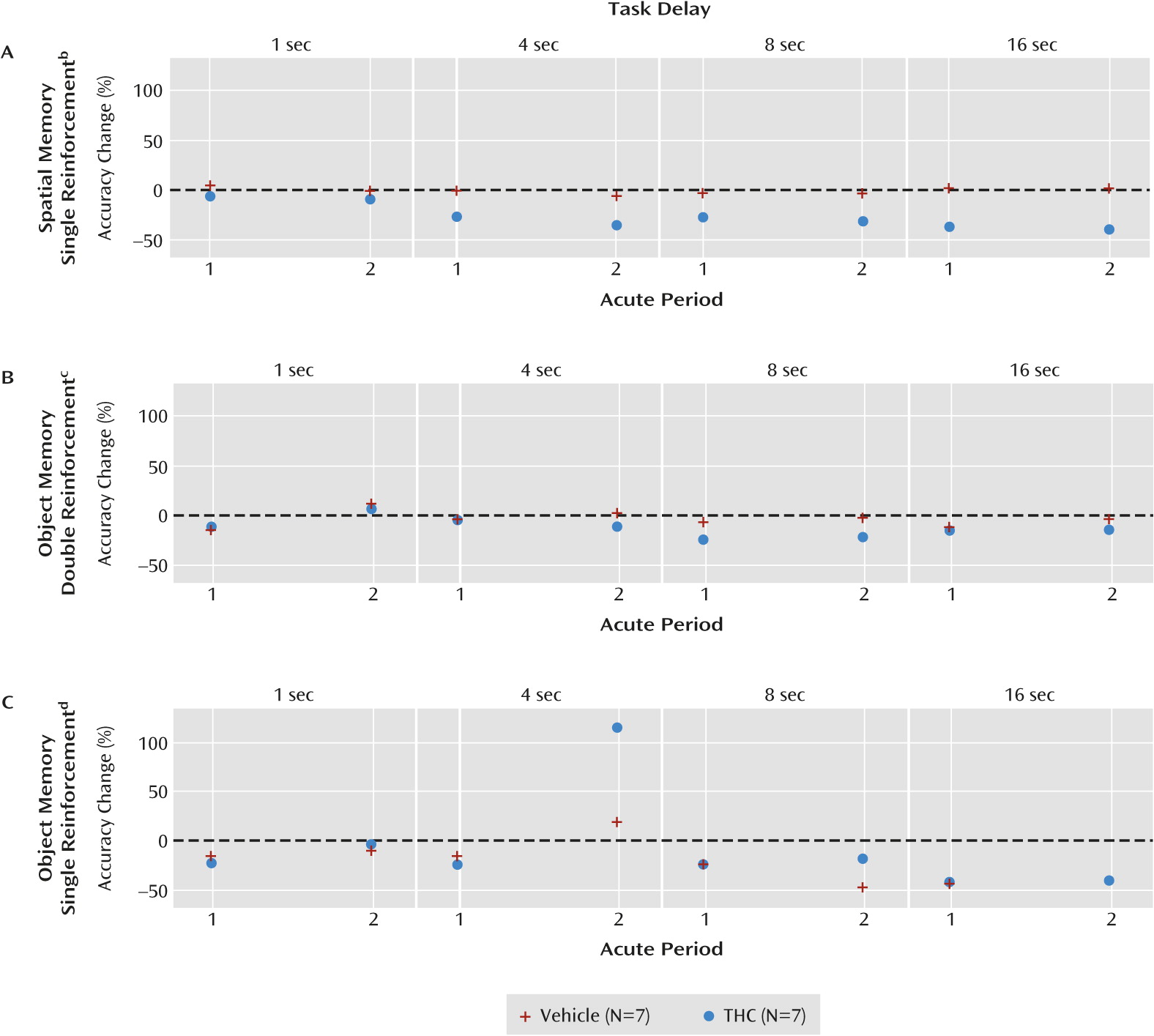
THC impaired spatial, but not object, working memory accuracy in a delay-dependent manner during the first acute period (
21). This same pattern was observed during the second acute period (
Figure 5 parts A, B, and C), and the spatial working memory impairments did not differ significantly between the two acute periods (group-by-period interaction: F=0.22, df=1, 70.9, p=0.64; group-by-period-by-delay interaction: F=0.07, df=3, 69.8, p=0.97). Consistent with a delay-dependent effect of THC on spatial working memory, the combined results from the two acute periods revealed an overall significant group-by-delay interaction (F=2.73, df=3, 70.1, p=0.05).