Schizophrenia is a heterogeneous disorder associated with complex emotional disturbances that are known to negatively interfere with patients’ functioning and quality of life (
1,
2). Psychopathological research has shown that emotional disturbances in schizophrenia encompass anxiety, depression, guilt, and irritability; conversely, blunting of affect has been observed in a subgroup of patients (
1,
2). In recent decades, several laboratory studies have investigated the nature of emotional disturbances in schizophrenia. Using facial emotional expression paradigms, it has been shown that facial emotion recognition is significantly impaired in schizophrenia patients (
3). Several studies have also investigated emotion experience in schizophrenia using emotion induction paradigms (e.g., film clips, affective pictures, and faces). These studies have shown that relative to control subjects, schizophrenia patients report greater negative emotions (aversion) and greater arousal levels in response to neutral stimuli (for meta-analyses, see references
4,
5). Echoing these results, studies using ecological momentary assessment or the experience sampling method have shown that schizophrenia patients consistently report more negative emotion than healthy volunteers in their daily lives (
6).
The importance, range, and functional significance of emotional disturbances in schizophrenia have led investigators to conduct several functional neuroimaging studies on the topic. Despite the heterogeneity of findings, meta-analyses of these studies have shown that schizophrenia patients display reduced activations in limbic regions, such as the amygdala and the parahippocampal gyrus, during the processing of negative emotional stimuli (
7–
9). Importantly, the main results of these meta-analyses seem to be observed in studies using affective faces or pictures, as well as studies using implicit (e.g., gender discrimination) or explicit (e.g., emotion recognition) task instructions. These neuroimaging results have been interpreted in light of the well-known blunting of affect and/or impaired emotion recognition abilities associated with schizophrenia.
One factor that has received insufficient attention in the field is that the reduced limbic activations observed during emotion processing in schizophrenia may be explained by the brain activity observed during the processing of neutral stimuli (e.g., control condition). In 2010, Anticevic et al. (
10) performed a seminal meta-analysis of 35 functional neuroimaging studies on emotion processing in schizophrenia that used the amygdala as a region of interest because of its well-known role in fear processing (
11). As observed by other research teams, the meta-analysis showed small reductions in amygdala activity in schizophrenia patients relative to healthy control subjects during emotion processing. More importantly, a subanalysis showed that this reduced amygdala activity is only present in studies examining the “negative emotion minus neutral” contrast. In the studies examining the “negative emotion only” condition, there were no significant differences in amygdala activations between schizophrenia patients and control subjects. This latter finding indirectly raised the possibility that schizophrenia patients actually display hyperactivations in response to neutral stimuli, which could explain (at least partially) the limbic hypoactivations that are usually reported during negative emotion processing in this population. In theory, it has been proposed that such aberrant amygdala reactivity to nonthreatening stimuli could fuel paranoid ideation in schizophrenia (
12). Consistent with this idea, a recent study using arterial spin labeling showed that amygdala activity at rest is elevated in patients with paranoid schizophrenia (
13). Likewise, positron emission tomography studies have shown increased tonic amygdala activity in schizophrenia patients regardless of experimental conditions (
14,
15).
Our objectives in this study were twofold. First, we aimed to perform a meta-analysis of functional neuroimaging studies examining the brain reactivity to neutral stimuli in schizophrenia patients while taking into account moderator variables such as psychotic symptoms, antipsychotic dosage, experimental stimuli, task instructions, and neuroimaging acquisition and processing parameters. Second, in order to test the aberrant salience hypothesis, we aimed to assess the brain reactivity of schizophrenia patients in response to emotionally negative stimuli in comparison to neutral stimuli. We hypothesized that schizophrenia patients would show increased activity in limbic regions such as the amygdala in response to emotionally neutral material as well as reduced or normal activity in response to emotionally negative stimuli.
Results
Twenty-three studies met inclusion criteria for the meta-analysis (
12,
16–
18,
25–
43) (
Figure 1). The study characteristics are summarized in
Table 1. The studies included 474 individuals with schizophrenia spectrum disorders who were compared with 472 healthy control subjects. The mean age of the schizophrenia patients was 35.3 years, and 70.8% of the sample were male. Patients had a mean antipsychotic dosage of 510.11 mg/day of chlorpromazine equivalents, and they had a mean score of 14.1 on the positive symptom subscale of the PANSS. Of the 23 studies, 14 used a whole-brain methodology and 17 used a smoothing kernel size of ≤8 mm. Eleven studies used an explicit emotional task, and 12 used only facial stimuli. Further details on the tasks, stimuli, and experimental contrasts used in the included studies are presented in Table S2 in the
online supplement.
Among the 23 studies (
12,
16–
18,
25–
43), no residual heterogeneity was observed between studies reporting increased activations (τ=0.0001, Q=11.57, df=11, p=0.397) or between studies reporting decreased activations (τ=0.034, Q=13.77, df=11, p=0.245) in schizophrenia patients compared with healthy subjects during emotionally neutral stimuli.
Responses to Emotionally Neutral Stimuli in Patients With Schizophrenia
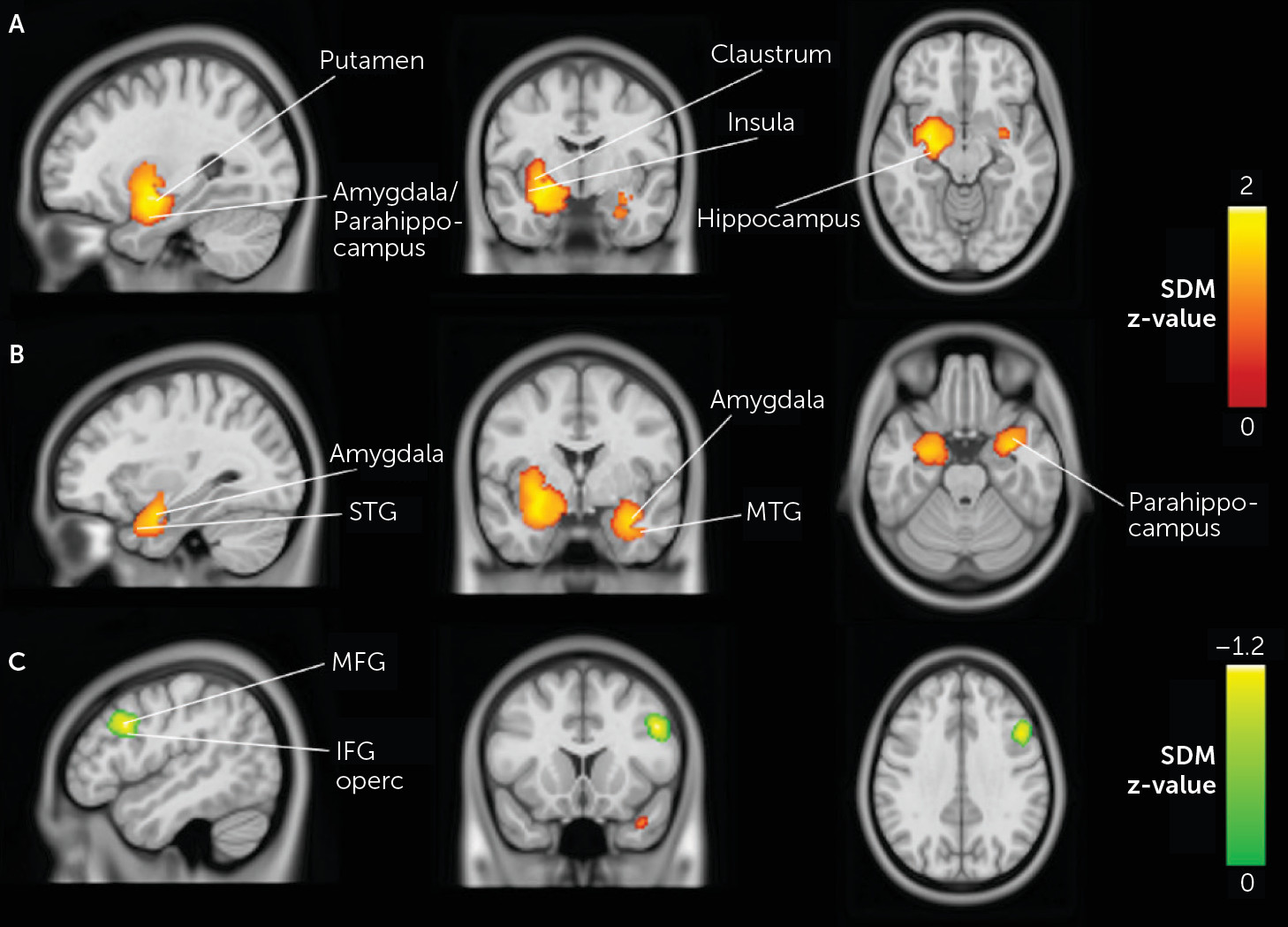
During neutral stimuli, schizophrenia patients showed significantly increased activations relative to healthy control subjects in large and robust limbic clusters bilaterally. Voxels from the left limbic cluster encompassed the putamen, the hippocampus, the amygdala, and the insula as well as the parahippocampal gyrus and the superior temporal gyrus (Z=1.90, cluster size=1,787, p<0.001). The right limbic cluster included the amygdala, the parahippocampus, and the middle temporal gyrus (Z=1.57, cluster size=776, p<0.001). Moreover, schizophrenia patients showed reduced activations, in comparison to healthy subjects, in the right middle frontal gyrus (Z=−1.16, cluster size=354, p<0.001), extending to the inferior frontal gyrus and the precentral gyrus (see
Table 2 and
Figure 2).
As shown by the jackknife analyses (see Table S3 in the
online supplement), the bilateral limbic clusters were highly replicable. Moreover, funnel plots and the Egger’s test suggested that these results were unlikely to be driven by small or noisy studies or publication bias (left peak: p=0.330; right peak: p=0.885). However, the result from the right inferior frontal gyrus was less replicable, as shown by the jackknife analyses (see Table S3 in the
online supplement). In fact, when the Ferri et al. study (
25) or the Ursu et al. study (
42) were excluded from the analysis, no significant peak was observed in this region. Although Egger’s test was nonsignificant (p=0.776), the funnel plot suggested that the effect observed in this particular region may have been driven by these two studies.
Subanalyses of emotionally neutral stimuli in patients with schizophrenia.
To enhance our understanding of the brain responses to emotionally neutral stimuli in individuals with schizophrenia, we performed several subanalyses.
First, when we restricted analyses to studies using only whole-brain analyses (N=14), we obtained results similar to those reported above: increased activations were observed in the schizophrenia group in the left putamen, left amygdala, left hippocampus, and left insula. However, we also observed a small but significant increase in activation in the left postcentral gyrus (Z=1.05, cluster size=212, p=0.0012) and decreased activation in the right fusiform gyrus (Z=−1.15, cluster size=300, p=0.0006) in schizophrenia patients in comparison to healthy subjects (see Table S4 in the online supplement). Conversely, in region-of-interest studies, we observed increased activations in the right amygdala (Z=1.65, cluster size=636, p=0.0004), encompassing the right parahippocampus, right middle temporal gyrus (Z=1.65, cluster size=424, p=0.0004), and left parahippocampal gyrus (Z=1.59, cluster size=269, p=0.0006), and decreased activations in right precentral gyrus (Z=−1.22, cluster size=363, p=0.0005), in schizophrenia patients compared with healthy subjects (see Table S5 in the online supplement).
Second, when we restricted the analyses to studies using facial stimuli (N=12), we also replicated the results on bilateral limbic activations as reported above (e.g., left and right putamen, left amygdala, left hippocampus, left parahippocampus, and left and right insula). Also, we observed a small but significant decreased activation in the left lingual gyrus (Z=−1.06, cluster size=142, p<0.001) (see Table S6 in the online supplement). Conversely, studies using stimuli other than facial expressions reported increased activations in the right angular gyrus (Z=1.13, cluster size=309, p=0.0006) and right amygdala (Z=1.06, cluster size=183, p=0.0009), as well as decreased activation in the opercular part of the right inferior frontal gyrus (Z=−1.72, cluster size=1072, p=0.00007), in patients compared with healthy control subjects (see Table S7 in the online supplement).
Third, when we restricted the analyses to studies using an explicit emotional task (N=11), we observed increased activation in the left putamen (Z=1.29, cluster size=601, p=0.00014) and decreased activation in the right fusiform gyrus (Z=−1.29, cluster size=688, p=0.0003) in schizophrenia patients in comparison to healthy subjects (see Table S8 in the online supplement). Conversely, studies using implicit task instructions (N=12) reported increased activations in the right uncus (Z=1.85, cluster size=1529, p=0.00004), encompassing the right amygdala and right parahippocampus, the left amygdala (Z=2.33, cluster size=1181, p<0.000001), encompassing the left hippocampus and left parahippocampus, as well as decreased activation in the right precentral gyrus (Z=−1.32, cluster size=976, p=0.0003) (see Table S9 in the online supplement).
Fourth, since there were too few studies using larger smoothing size kernel (N=6), no further subanalyses were performed. However, when we performed a subanalysis within studies using a smoothing kernel of ≤8 mm (N=17), we observed results similar to those from the main analysis (e.g., left putamen, left and right amygdala, left hippocampus, left and right parahippocampus, and left insula) (see Table S10 in the online supplement). Finally, meta-regressions revealed no significant relationships between results from the main analysis and score on the positive symptom subscale of the PANSS (p values, 0.443–0.970), antipsychotic dosage in chlorpromazine equivalents (p values, 0.495–0.914), and repetition time of functional volumes (p values, 0.303–0.515).
Responses to Emotionally Negative Versus Neutral Stimuli in Schizophrenia Patients
Of the 23 studies included in the meta-analysis, 16 reported responses to emotionally negative stimuli versus neutral stimuli in schizophrenia patients compared with healthy control subjects (for detailed information about the studies, see Table S11 in the online supplement).
No residual heterogeneity was observed between studies (positive peaks: τ=0.075, Q=7.35, df=5, p=0.196; negative peaks: τ=0.043, Q=4.28, df=3, p=0.233). Results of the meta-analysis (N=16) revealed that there were no significant between-group differences in activations between schizophrenia patients and healthy subjects in response to emotionally negative stimuli.
Discussion
To our knowledge, this is the first functional neuroimaging meta-analysis on the processing of emotionally neutral material in schizophrenia. Whereas the meta-analysis of Anticevic et al. (
10) inferred that the amygdala may be overactivated in schizophrenia during the neutral condition, here we were able to directly test this hypothesis using data from 23 functional neuroimaging studies. Results showed increased activations in schizophrenia patients relative to healthy control subjects in the left and right amygdala, the left putamen, the left hippocampus, the left and right parahippocampal gyrus, and the left insula. Conversely, reduced activations were observed in the right middle frontal gyrus, a region involved in emotion regulation (
44). Importantly, no differences in brain activity were observed between groups when examining the emotion minus neutral contrast. Moreover, the limbic results were homogeneous and robust. In the case of the middle frontal gyrus, results were mostly driven by the influence of only two studies. Subanalyses and meta-regression analyses revealed no noticeable impact on results of temporal resolution and smoothing level. On the other hand, the type of stimuli (e.g., faces) and the type of task instruction (e.g., explicit processing) had a significant impact on results, as the most widespread limbic hyperactivations in schizophrenia patients were observed in studies using facial stimuli and studies using implicit task instructions having a lower cognitive load (
45). Finally, the increased activations observed in the right amygdala and the left and right parahippocampal gyrus were mostly driven by studies adopting a region-of-interest approach, since no differences between schizophrenia patients and control subjects were observed in these two brain regions in the set of studies that performed whole-brain analyses.
The most important finding of this meta-analysis is the confirmation of the hypothesis of increased activity of the left amygdala specifically in response to emotionally neutral stimuli in schizophrenia patients. Given the key role of the amygdala in threat detection (
11), this result strongly suggests that schizophrenia patients confer aberrant emotional significance to nonthreatening stimuli. In theory, this neurophysiologic process could fuel delusional thinking in schizophrenia. Another contribution of this meta-analysis is that it showed that other important subcortical regions (e.g., putamen, hippocampus, insula) are also more activated in schizophrenia patients, apart from the amygdala. The finding of increased activity in the left putamen in schizophrenia patients in our meta-analysis is consistent with the dopaminergic hypothesis of psychosis. Using PET imaging, several studies have indeed shown that striatal dopamine function is increased in schizophrenia and, more precisely, that dopaminergic alterations are more prominent in the associative rather than the ventral striatum (
46). As for the (left) hippocampus, this region is well known for playing a key role in memory formation and associative learning (
47). The increased activations observed in this region in schizophrenia during the viewing of emotionally neutral stimuli may therefore be interpreted as a difficulty in making relevant associations between emotionally neutral stimuli and their context. As for the (left) insula, it plays a key role in interoceptive awareness, and as such, the increased activations observed in this region suggest that irrelevant stimuli are inappropriately associated with somatic changes in schizophrenia. Taken together with the normal activity observed in schizophrenia during emotion processing, these complex neuroimaging results are clearly consistent with the aberrant salience hypothesis, which proposes that psychosis arises from an aberrant attribution of motivational or emotional significance to irrelevant stimuli that are out of context (
48). In this model, delusions arise from a top-down cognitive effort of the patient to make sense of the aberrant salience experiences. An indirect implication of the model is that aberrant salience experiences may give rise to psychiatric symptoms other than delusions, depending on how these experiences are apprehended. In that regard, no study has examined the relationship between the aberrant limbic reactivity to neutral stimuli and anxious-depressive symptoms in schizophrenia. However, preliminary fMRI results have shown that the limbic reactivity to emotionally neutral faces is increased in mood and anxiety disorders (
49).
To better understand the meaning of the increased limbic reactivity observed in schizophrenia during the processing of nonthreatening stimuli, the use of more sophisticated neuroimaging analyses, such as multivoxel pattern analyses or independent component analyses, would be required. Perhaps more importantly, careful attention will need to be paid to the experimental paradigms used to study this phenomenon. In that regard, two particular avenues seem promising. As the studies included in our meta-analysis all included an emotional condition and a neutral condition, a first possibility is that neutral stimuli may acquire aberrant significance in schizophrenia via impaired associative learning. This is a likely possibility given that we found increased activations in the hippocampus. For instance, in one of the studies included in the meta-analysis, the investigators used an aversive Pavlovian conditioning paradigm and showed increased activations in schizophrenia patients in limbic regions (e.g., ventral striatum and hippocampus) in response to conditioned stimuli paired to unconditioned neutral stimuli (
27). Another relevant possibility has to do with the growing body of work in social neuroscience showing that faces that are neutral in terms of expressed emotion are not necessarily neutral in every other aspect. Indeed, emotionally neutral faces convey subtle information (e.g., physiognomic features) implicitly processed by the brain. When watching neutral faces, people make several spontaneous judgments (implicit or explicit) about the face’s level of attractiveness, introversion/extraversion, and trustworthiness. In the present meta-analysis, the most prominent limbic effects were observed in studies using facial stimuli. In theory, schizophrenia patients may underestimate the trustworthiness of others’ faces, and this bias could fuel suspicious thoughts. Interestingly, preliminary fMRI studies have shown limbic alterations in schizophrenia patients who were asked to make trustworthiness judgments about emotionally neutral faces (
13). Despite the promise of this social neuroscience approach, it must be remembered that the physiognomic features of faces judged as being untrustworthy are not entirely independent from the physiognomic features of faces expressing anger (
50).
In the past decade or so, preliminary studies have begun to pay attention to brain reactivity to neutral stimuli in individuals at risk for psychosis. Notably, a neuroimaging study examining emotion processing in youths at risk for psychosis (
51) showed that the aberrant limbic reactivity to neutral stimuli (e.g., dynamic faces) is detectable as early as age 14. However, some studies have failed to observe this effect (regardless of the brain region) (
52,
53). One of the critical factors that may compromise the acquisition of consistent findings here is the wide range of definitions of psychosis risk. Future studies will need to pay greater attention to this population using well-defined inclusion criteria.
Our study has some limitations that need to be acknowledged. First, some studies included in the meta-analysis used a region-of-interest approach, and used the amygdala as the seed region, for instance. The choice of this approach may have inflated results in the chosen regions of interest and underestimated effects outside these regions. Because of this concern, we performed a subanalysis and found that only the results regarding the right amygdala and the left and right parahippocampal gyrus were driven by the region-of-interest approach. The effects seen in the left amygdala, the left putamen, the left hippocampus, and the left insula were not confounded by this factor.
Another limitation of our meta-analysis is that the vast majority of patients were on antipsychotic treatment when they underwent scanning. However, antipsychotics all block dopamine D
2 receptors and are thought to dampen aberrant salience, not to stimulate it (
46). Moreover, we performed a meta-regression analysis using antipsychotic dosage in chlorpromazine equivalents and found no association with any of our main results.
A third limitation of our meta-analysis is that the results may have been influenced by publication bias. Since it is not common to report activations during the neutral condition, it is possible that investigators who decided to do so were those who found significant differences between patients and control subjects. Aware of this possibility, we performed analyses of publication bias and found that the results regarding the middle frontal gyrus were driven by a minority of studies. Importantly, however, none of the findings related to the limbic system (e.g., amygdala, putamen, hippocampus, parahippocampal gyrus, and insula) were influenced by publication bias.
A fourth limitation of the meta-analysis is that the subanalysis on negative emotions was based on the 16 studies (of the 23 studies in the analysis) that reported relevant data. In previous meta-analyses that had a primary objective of examining negative emotions, more studies were included, and their aggregation showed reduced limbic activations during the processing of negative emotions in schizophrenia (
7,
10).
Finally, no correlation was found between the limbic regions observed to be more activated in schizophrenia patients in response to neutral stimuli and the positive symptoms of the disorder. If the increased brain reactivity to neutral stimuli is involved in the generation of paranoid or delusional ideas, we would have expected to observe a correlation across studies. One factor that may have compromised our ability to detect this expected association is that the meta-regression analysis was performed with the PANSS positive symptom score, which measures four symptoms (e.g., hallucinations, disorganization, hostility, excitement) other than delusional thinking. Moreover, the mean PANSS positive symptom score was rather low across studies.