Early adversity (e.g., childhood maltreatment) is associated with significantly elevated risk for negative developmental outcomes, including psychopathology and atypical neurobiological development (
1,
2). One likely pathway through which early adversity may confer heightened risk for these problems is attachment difficulties (
3). In particular, it has been well documented that children who experience maltreatment are less likely to develop secure attachments to parents than children who are not maltreated (
3,
4). Insecure or disorganized attachments, in turn, place children at greater risk for internalizing and externalizing behavioral problems than secure attachments (
5,
6). Findings from both human and animal investigations have identified neural responses to parent cues during childhood as a candidate biological mechanism linking early caregiving experiences to attachment-related processes and mental health outcomes (
7–
12). However, although work with animals has established the causal role of early parenting experiences in offspring reactivity to parent cues (
12), such work with humans has been largely correlational (
7–
11). In the present study, we leveraged a randomized clinical trial to test the causal impact of an early parenting intervention on the neural processing of parent cues and on psychosocial functioning in children.
Caregiving that is attuned to the needs of the offspring is critical for the development of neural systems underlying social functioning in altricial mammals, including humans (
10). In caring for offspring, mammalian mothers utilize a distinct subcortical network supporting maternal behavior (
13), which in humans evolved to include cortical regions—such as the medial prefrontal cortex (mPFC), anterior cingulate cortex (ACC), dorsolateral PFC, insula, inferior frontal cortex, orbitofrontal cortex (OFC), and temporoparietal junction (
14)—that are implicated in social cognition, emotion regulation, and behavior. Relatively less is known about how children respond to attachment cues at the level of brain activation; however, recent findings indicate significant overlap with parents’ brain responses. For example, when children listened to their own mother’s voice, compared with when they listened to female control voices, they exhibited greater activation in a host of brain regions, including the mPFC, ACC, insula, OFC, precuneus, posterior cingulate cortex (PCC), angular gyrus, fusiform gyrus, and amygdala, with between-area connectivity during presentation of mothers’ voices predicting children’s social communication skills (
11). This pattern of activation is not specific to auditory maternal cues, because many of the same brain areas (e.g., the mPFC, PCC, fusiform gyrus, and amygdala) have shown greater activation among children and adolescents while they were viewing pictures of their own mothers compared with viewing pictures of female control subjects (
7–
9). An important remaining question is that of what factors contribute to the development of children’s responses (i.e., how these patterns of activation are transmitted across generations).
Perhaps unsurprisingly, evidence thus far points toward sensitive parent-child interaction as critical to the development of typical attachment cue processing. For example, mother-child social synchrony during play has been associated with children’s cortical (i.e., the fusiform gyrus, superior temporal sulcus, and insula) theta and gamma oscillatory activity measured by magnetoencephalography in response to viewing themselves interact with their own mothers compared with viewing unfamiliar mother-child interactions (
10). Moreover, children exposed to early caregiver deprivation (i.e., those with a history of previous institutional care) show less left amygdala discrimination between mother and stranger stimuli than children with no history of institutional care, with less amygdala discrimination being associated with older age at adoption and greater indiscriminate friendliness (indicating atypical attachment development) (
8). Together, these findings suggest that brain areas implicated in social cognition and emotion processing are influenced by parenting and early adversity and that they may play a role in psychosocial outcomes, in line with the general view of attachment theory that children’s social representations of attachment figures influence their internal working models of the world (
15). Importantly, however, most such studies to date have been correlational in nature and do not permit causal interpretations with regard to potential effects on brain development and subsequent behavior.
Early interventions that enhance parenting quality have been shown to improve atypical developmental trajectories associated with early adversity (
16). Specifically, early parenting interventions can enhance parental responsiveness (
4,
17,
18) and improve infants’ attachment quality (
4,
17–
19) and physiological and behavioral regulation (
20,
21). However, the neural mechanisms through which these early interventions improve psychosocial outcomes remain poorly understood.
The Attachment and Biobehavioral Catch-Up (ABC) intervention (
22) is a well-characterized, evidence-based early parenting intervention that may permit researchers to investigate how children’s underlying neurobiology changes in response to early intervention delivered to their parents. ABC is delivered across 10 in-home sessions by trained parent coaches and has been shown to be efficacious in improving parent and child outcomes through multiple randomized controlled trials involving several vulnerable populations, including children in the foster care system, children living with birth parents following involvement with Child Protective Services (CPS), and children who were adopted internationally. The intervention aims to increase rates of secure attachment (as well as reduce rates of disorganized attachment) and improve children’s behavioral and biological regulation by increasing parental nurturance when children are distressed, increasing parental sensitivity and positive regard when children are not distressed, and decreasing frightening and intrusive parental behavior. Parents randomly assigned to receive the ABC intervention have demonstrated greater sensitivity and positive regard, as well as lower intrusiveness and withdrawal, than parents randomly assigned to receive a control intervention (
23). The effects go beyond the parents: children whose parents received the ABC intervention demonstrated more adaptive patterns of autonomic regulation (
24), more normative diurnal cortisol rhythms (
20,
21), greater executive functioning skills (
25,
26), stronger emotion regulation skills (
27), and decreased disorganized attachment (
23) than children of parents who received control interventions.
Given the centrality of parental influence in fostering the social brain and behavioral development, we aimed to test, in a randomized clinical trial, the causal impact of an early parenting intervention (ABC) on children’s neural processing of parent cues and children’s psychosocial functioning in middle childhood. We hypothesized that compared with high-risk children whose parents received a control intervention, high-risk children whose parents received the ABC intervention would show greater neural responsivity to parent cues in cortical regions implicated in social and emotional processing and would exhibit better psychosocial functioning as measured by parent report. In addition, if there were group differences in neural responsivity to parent cues, a secondary aim was to test whether such group differences mediated differences in children’s psychosocial functioning.
Methods
Participants
Families (N=212) with infant children were originally recruited as part of a randomized clinical trial in a major U.S. Mid-Atlantic city. As part of a citywide initiative designed to redirect children from foster care, families were referred from CPS because of the risk for abuse or neglect. At the time of recruitment, enrolled families were randomly assigned to receive either the ABC intervention or a control intervention (described below). Families were not informed about their intervention group assignments. At the preintervention assessment, children across the intervention groups did not differ significantly in age, race, or diurnal cortisol levels (
20), and caregivers did not differ significantly in age, educational attainment, race (
19), parental sensitivity, or attachment-related representations (
24). Of the 212 families enrolled in the randomized controlled trial, 183 participated in initial postintervention follow-up assessments, and 112 participated in 8-year follow-up assessments (see the flow diagram in the
online supplement). A subset of families who participated in the 8-year follow-up assessments were invited to participate in the present functional MRI (fMRI) substudy. To maximize the chances of successful scans, children who successfully completed an EEG assessment as part of an 8-year follow-up visit were subsequently invited to participate in this fMRI substudy. Ultimately, 54 high-risk children (ABC [experimental] intervention: N=27; Developmental Education for Families [DEF] [control] intervention: N=27) ages 8.1–12.1 years participated in this fMRI substudy (for demographic characteristics, see Table S1 in the
online supplement).
For comparison with the two high-risk groups (the ABC intervention group and the control intervention group), a new sample of 83 non-CPS-referred children who did not receive any intervention was recruited at age 8 through local community centers and schools. This low-risk sample was matched to the CPS-referred sample on race and gender. Families were ineligible for recruitment to the low-risk sample if they had a history of CPS involvement. Similar to the high-risk sample, comparison children who completed the 8-year EEG assessment were subsequently invited to participate in this fMRI substudy. The fMRI low-risk comparison sample comprised 26 children ages 9.1–11.0 years. Recruitment for the fMRI substudy ended after a total of 80 children had participated in the substudy (ABC intervention group: N=27; DEF intervention group: N=27; low-risk group: N=26).
Experimental intervention.
ABC is a brief (10-session) home-based parenting intervention that promotes sensitive caregiving. This intervention focuses on three main behavioral targets for parents: increasing sensitivity to child signals, increasing nurturance to child distress, and decreasing frightening and harsh behaviors. In addition to manualized content, intervention sessions consist of parent coaches providing in-the-moment commenting and feedback to support parents in identifying their children’s signals and providing responsive care (
22).
Control intervention.
DEF is an adaptation of existing interventions (e.g.,
28) that have been shown to promote development of children’s motor skills, cognition, and language abilities. Components of the intervention related to parental sensitivity were removed for this substudy to help distinguish it from ABC.
Procedure
As noted above, families enrolled in the larger longitudinal study investigating the efficacy of the ABC intervention were invited to participate in this fMRI substudy. After parents provided informed consent and children provided assent, children were acclimated to the scanner using an MRI replica before the scanning session, which typically occurred within 2 weeks of the practice session. The protocol was approved by the institutional review board at the University of Delaware.
Questionnaires
Child Behavior Checklist (CBCL).
Parents completed the CBCL for ages 6–18 (
29) in the laboratory as part of a battery of measures. The CBCL asks about 113 emotional and behavioral problems rated from 0 (not true) to 2 (very true or often true). In this study, two items related to suicidality and self-harm were removed from the questionnaire. Raw total scores were used in analyses as a measure of psychosocial functioning. In this study, the CBCL had excellent internal consistency (alpha=0.94).
Kerns Security Scale.
Children completed the Kerns Security Scale (
30), which was used to measure their perceived security to their mothers. The scale consists of 15 items divided into three subscales mapping onto parent responsivity or availability, reliability during times of stress, and interest in communicating with the parent. Higher scores indicate greater feelings of security in the mother-child relationship. The three subscales were collapsed for analyses. In this study, the scale had moderate internal consistency (alpha=0.68), possibly as a result of the relatively small number of items.
Imaging
Parent-stranger fMRI task.
In the scanner, children completed a parent-stranger task (
8,
9) in which they were presented with eight alternating blocks (28 seconds each, for a total task time of 4 minutes and 54 seconds) of color photographs of their own mother or another child’s mother (i.e., a stranger matched to the parent on ethnicity, age, and body type) exhibiting smiling and neutral facial expressions. Only mother-child dyads were included in this study, and thus the parent and stranger sex was always female. Each alternating block consisted of either images of the child’s parent or images of the stranger. To ensure attention to the task, participants were instructed to respond with a button press to only the smiling stimuli; however, smiling and neutral trials were collapsed together for analyses. Additionally, as a result of the nature of a block design, analyses of each emotion were not possible.
Image acquisition.
Images were acquired with a Siemens Prisma 3-T MRI scanner (Siemens, Erlangen, Germany). A whole-brain, high-resolution, T1-weighted anatomical scan (magnetization-prepared rapid gradient-echo; in-plane resolution, 256×256; field of view, 256 mm; sagittal slices, 192×1 mm) was used for transformation and localization of each participant’s functional data into Montreal Neurological Institute 152 (MNI 152) space. For the parent-stranger functional task, T2*-weighted echo-planar images (34 slices) were acquired using an oblique angle of approximately 30° from each participant’s position (4-mm slice thickness [skip=0], repetition time, 2,000 ms; echo time, 30 ms; flip angle, 90°; matrix, 64×64).
fMRI preprocessing.
Functional imaging data were preprocessed and analyzed using FMRIB Software Library (FSL), version 6.0.1 (
31). Preprocessing, single-subject statistics, and higher-level analyses were performed using the FSL fMRI Expert Analysis tool (
32). Preprocessing steps included slice-timing correction, motion correction (with FMRIB’s linear registration tool [
33]), image registration to the first volume, smoothing with an anisotropic 6-mm Gaussian kernel (full width at half maximum), time series normalization, and transformation into MNI 152 space. Eight explanatory variables were included in the regression model (six motion parameters and the two stimulus types [mother and stranger]). Volumes with excessive frame-wise motion (>0.9 mm from the adjacent volume) were censored (
34), and participants with >30% total volumes censored were excluded from analysis. From the low-risk group, one participant was excluded because of excessive head motion, two were excluded because of image registration problems, and one did not complete the parent-stranger task (for details on exclusion criteria for the high-risk group, see the flow diagram in the
online supplement). The final substudy sample comprised 68 children for analyses (ABC, N=22; DEF, N=24; low-risk, N=22). In this final sample, there were no significant group differences in age (F=0.602, df=2, 65, p=0.551) or sex (χ
2=0.123, df=2, p=0.940).
Statistical Analysis
Whole-brain analyses were performed to test the within-subject effect of stimulus type (mother compared with stranger) on activity in cortical and subcortical brain regions, as well as possible group differences in this stimulus effect in a series of planned comparisons. The FSL FLAME 1 mixed-effects model was used with automatic outlier de-weighting. Clusters of blood-oxygen-level-dependent (BOLD) activation were considered significant if the Z value was >2.3 with a corrected cluster significance threshold of 0.05. In addition, given the number of group comparisons, the family-wise error rate was further controlled using FSL’s “randomise” function with threshold-free cluster enhancement. Brain structure labels were estimated probabilistically with the Harvard-Oxford cortical and subcortical structural atlases in FSL using the “autoaq” function. Lastly, causal mediation analysis (
35) was performed in R, version 3.6.1, using the “mediation” package (
36) to determine whether intervention group differences in mother-specific BOLD reactivity mediated the relationship between intervention group assignment and psychosocial outcomes.
Results
Behavior
To test the main effects and interactions of stimulus type and group, 2×3 (stimulus type: mother, stranger; and group: ABC, DEF, low-risk) analyses of variance were performed for hit rate, false alarm rate, hit reaction time, and false alarm reaction time. There were no significant main or interaction effects for any of these behavioral measures (all p values >0.05). These effects remained nonsignificant when controlling for child age and child sex (all p values >0.05).
Imaging
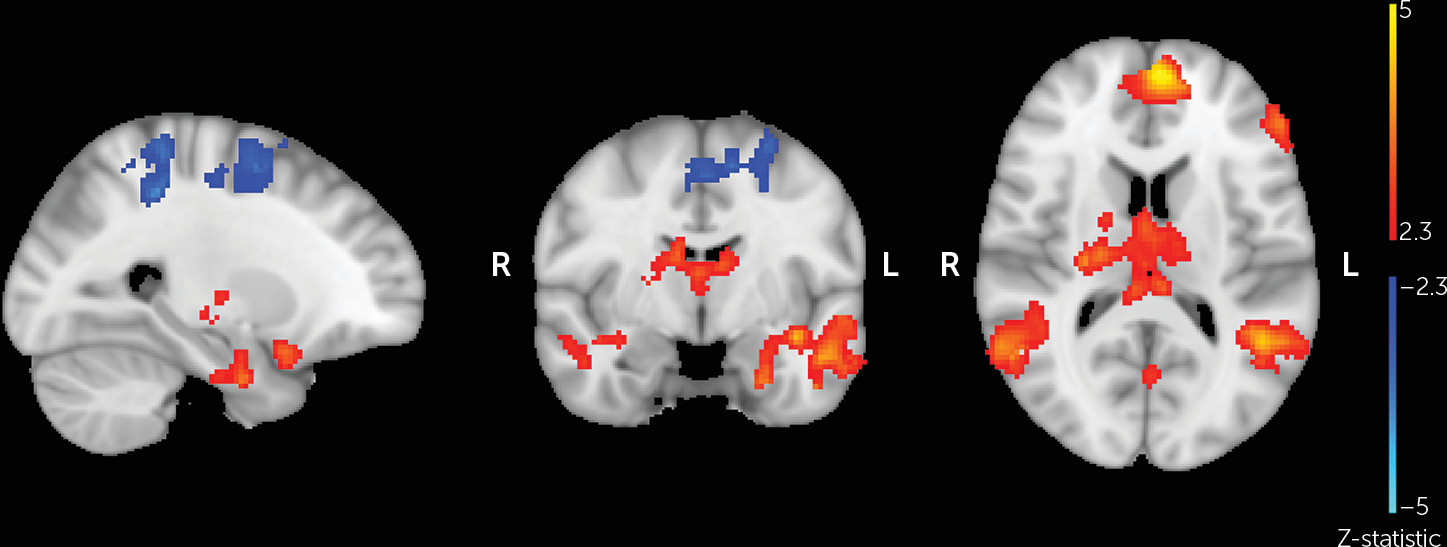
In order to verify that the parent-stranger task elicited the expected neural responses, whole-brain analysis was performed comparing parent and stranger trials across all participants (
Figure 1). Compared with viewing the stranger photographs, viewing pictures of one’s own mother was associated with greater activation in clusters including the left and right amygdala, hippocampus, thalamus, and OFC (p<0.025) (
Table 1). Conversely, viewing pictures of the stranger was associated with greater activation in clusters including the precentral and postcentral gyri (p<0.05) compared with viewing pictures of one’s own mother.
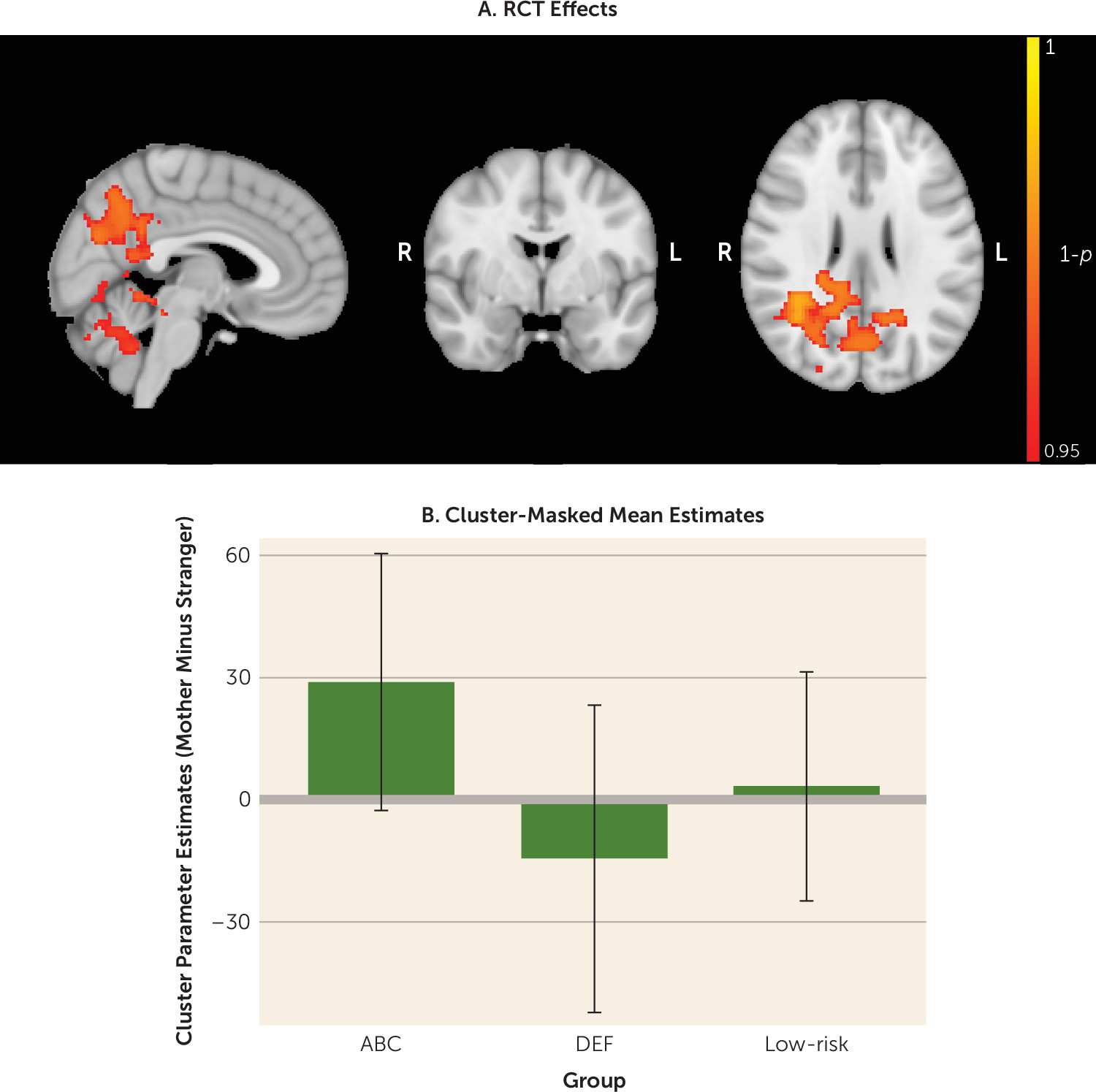
Whole-brain analysis comparing the three groups’ responsivity to the mothers’ faces compared with strangers’ faces revealed significant differences between the two high-risk groups. Specifically, children whose parents received the ABC intervention exhibited greater activation in response to mother (compared with stranger) images than children whose parents received the DEF intervention. These effects were observed in clusters including the precuneus and cuneal cortex, PCC, middle temporal gyrus, lateral occipital cortex, angular gyrus, and hippocampus (p<0.05, corrected) (
Table 2,
Figure 2). The observed intervention effects remained significant when controlling for child age and child sex (p<0.05, corrected). The ABC group also exhibited somewhat greater activation in response to the mother (compared with stranger) than the low-risk group in the left and right precuneus and left and right cingulate gyrus; however, this group difference did not survive correction for multiple group comparisons (p<0.001, uncorrected; p=0.421, corrected). There were no significant whole-brain group differences between the DEF and low-risk groups.
Additionally, on the basis of findings from previous studies (
8,
9), we performed amygdala region-of-interest analyses with separate left and right amygdala regions of interest based on the Harvard-Oxford Subcortical Structural Atlas. There were no significant group differences in the left amygdala (mother only: F=0.117, df=2, 65, p=0.890, η
2=0.004; stranger only: F=0.800, df=2, 65, p=0.454, η
2=0.024; mother compared with stranger: F=1.711, df=2, 65, p=0.189, η
2=0.050) or right amygdala (mother only: F=0.529, df=2, 65, p=0.592, η
2=0.016; stranger only: F=1.804, df=2, 65, p=0.173, η
2=0.053; mother compared with stranger: F=2.371, df=2, 65, p=0.101, η
2=0.068). These amygdala group effects remained nonsignificant when controlling for child age and child sex (all p values >0.05).
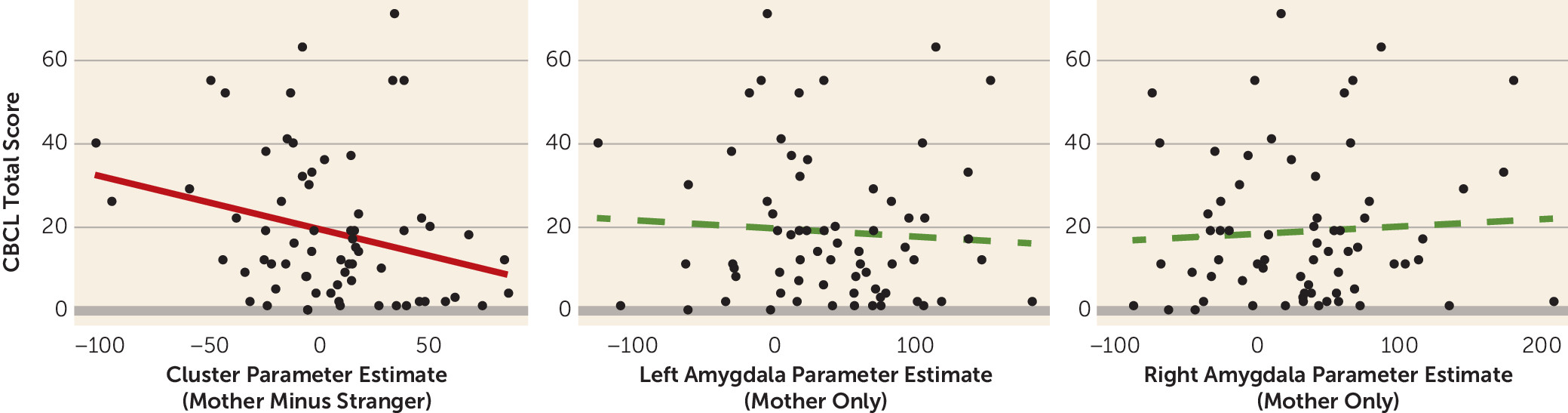
Additionally, there were no significant associations between task behavior and task-related BOLD activation in the left or right amygdala or in the clusters of brain regions that differentiated the ABC intervention and the DEF intervention (primarily the precuneus and cingulate gyrus [see above]) in any of the contrasts of interest (i.e., mother compared with stranger, mother only, stranger only; all p values >0.05).
Questionnaires
Descriptive statistics and correlations among questionnaire measures are presented in the tables in the
online supplement. There were no significant group differences in the CBCL total score (F=0.473, df=2, 64, p=0.625, η
2=0.015) or Kerns Security Scale total score (F=0.391, df=2, 65, p=0.678, η
2=0.012). Group effects on questionnaire measures remained nonsignificant when controlling for child age and child sex (all p values >0.05). The descriptive statistics of subscale scores are presented in Table S1 in the
online supplement. However, greater activity in the clusters differentiating the ABC and DEF groups (mother compared with stranger) was associated with lower CBCL total scores (r=−0.27, df=65, p=0.030) (
Figure 3), indicating that greater activation in response to viewing the mother (compared with stranger) in these areas was associated with fewer parent-reported behavior problems. Associations between Kerns Security Scale scores and task-related BOLD measures are presented in Table S2 in the
online supplement.
Mediation Analysis
In order to test for potential indirect effects of intervention group on psychosocial outcomes within the high-risk sample, causal mediation analysis (
35,
36) was performed with 10,000 permutations using intervention group assignment as the predictor, BOLD reactivity during the parent-stranger task as the mediator, and total CBCL score as the outcome. Specifically, the mediator consisted of the average beta weights from the mother-stranger contrast cluster that significantly differentiated the ABC and DEF groups. Although there was no significant direct effect of intervention on CBCL score (p>0.05), there was a significant indirect effect of intervention on CBCL score (average mediation estimate=−7.453, 95% CI=−16.773, −0.320, p=0.037). This estimate indicates the average decrease in total CBCL scores that was attributable to the effect of the ABC intervention on BOLD reactivity during the parent-stranger task. Additional mediation models involving CBCL subscale scores and Kerns Security Scale scores are presented in Table S3 in the
online supplement.
Discussion
In a randomized clinical trial, we examined the causal impact of an early intervention that enhances parenting on children’s neural processing of maternal cues and on their psychosocial functioning during middle childhood. Most previous work in this area has been correlational and thus vulnerable to numerous threats to internal validity. We hypothesized that compared with high-risk children of mothers randomly assigned to receive the control intervention, high-risk children of mothers randomly assigned to receive the ABC intervention would show greater neural responsivity to maternal cues in areas implicated in social processing (e.g., the amygdala and cortical regions such as the OFC, PCC, insula, temporal fusiform cortex, and precuneus cortex). When looking across both high-risk and low-risk children, consistent with previous child neuroimaging studies involving presentation of maternal cues (
9,
11), mother-specific activation was observed across a wide variety of brain regions, including frontal and sensory cortices as well as subcortical structures. With regard to this study’s main hypothesis, children whose parents received the ABC intervention exhibited greater responsivity to maternal cues in clusters of brain regions including the precuneus and cuneal cortex, PCC, middle temporal gyrus, temporal fusiform cortex, lateral occipital cortex, angular gyrus, hippocampus, and others. Children in the ABC group also exhibited somewhat greater responsivity to maternal cues in a subset of these brain regions (the precuneus and PCC) compared with children in the low-risk comparison group, but this effect did not survive correction for multiple group comparisons. Somewhat contrary to what was predicted, there were no significant group differences in amygdala activation; however, given that the amygdala is a relatively small structure with low magnetic resonance signal, it is possible that our study was underpowered to detect group differences in amygdala activation. Nevertheless, clear intervention effects were observed in the high-risk sample, allowing causal interpretations of the effects of a parenting intervention on neural reactivity to attachment cues among children. Results suggest that the ABC intervention may enhance children’s brain development despite the fact that it targets parental sensitivity rather than intervening with the child directly.
Many of the brain areas whose maternal cue-related activation was augmented by the ABC intervention are also implicated in aspects of social cognition, such as theory of mind, and other aspects of social representation (
37), suggesting that this intervention could enhance brain regions supporting children’s social cognitive development. Cortical midline structures, which include the precuneus and PCC, have been implicated in understanding “complex psychological aspects of others,” such as their attitudes (
38). The precuneus, in particular, is a major node of the cortical midline structures thought to be involved in elaborating highly integrated and associative information, such as maintaining self-other representations across multiple domains, and has direct connections to the mirror neuron system involved in imitative behavior and social cognition (
38). The mirror neuron system, which also includes regions whose maternal cue-related activity was augmented by ABC (e.g., the superior parietal lobule and inferior occipital cortex), is especially sensitive to self-other mappings, such that its level of activation tracks the degree of schematic overlap between the self and a perceived other (
39). Although based on reverse inference, these findings, when taken together, point to a possible interpretation wherein ABC enhances children’s relational representation of their mothers, resulting in heightened activation of cortical midline structures and mirror neuron system brain regions while viewing pictures of the caregiver. However, because the control (i.e., stranger) face was unfamiliar to the participant, further study is needed to determine whether the observed effect of the parenting intervention on facial processing is specific to maternal cues or generalizable to familiar others. This would help clarify the extent to which intervention effects are limited to reactivity to parent cues compared with having a more global impact on social processing.
Given that the ABC intervention has been associated with improvements in executive functioning and emotion regulation skills that endure through at least early childhood (
25–
27), we also hypothesized that children whose parents received this intervention might exhibit better psychosocial functioning as late as middle childhood compared with children whose parents received the control intervention (DEF). Although we did not find a significant direct effect of intervention on CBCL scores at the age of scanning (8.1–12.1 years), there was a significant indirect effect of intervention on total CBCL scores mediated through maternal-specific activation of the clusters of brain regions that significantly differentiated the ABC group from the control intervention group (e.g., the precuneus, PCC, superior parietal lobule, and inferior occipital cortex). A potential limitation of this mediation model is that because the mediator and outcome were measured at the same time point, temporal precedence of the mediator over the outcome cannot be established. However, because there was no significant direct effect of intervention group on CBCL scores, an alternative model in which the association between the intervention group and task-related BOLD reactivity is mediated by psychosocial functioning can be ruled out. If it is indeed the case that the mediator reflects enhancement of children’s parent-child relational representations, the results of the significant mediation model would be consistent with the view that attachment figures influence children’s internal working model of the social world, which in turn influences children’s psychosocial functioning (
15).
In addition to the limitations mentioned earlier, it should be noted that because of a lack of detailed CPS referral information, the high-risk group (comprising families who received the ABC or control intervention) combined children who were likely to have experienced neglect, abuse, or both. Although children with substantiated and unsubstantiated allegations of maltreatment are at similar risk for negative behavioral and developmental outcomes (
40), it is not unreasonable to suppose that an intervention aimed at enhancing parental sensitivity may have a differential effect as a function of the type of early adversity or maltreatment a child experienced; however, the fact that significant intervention effects emerged in a relatively small sample despite this potential heterogeneity highlights the value of early intervention.
Overall, the significant indirect effects of the ABC intervention revealed by mediation analysis suggest that in addition to the intervention causing greater mother-specific activation of the empirically identified brain regions (perhaps suggesting enhancement of the child’s parent-child relational representation), this pattern of activation may be indicative of improved parent-child relationship factors that are enhanced by this intervention and are associated with better psychosocial outcomes. In other words, these results suggest a possible neural pathway through which an early parenting intervention—in this case, ABC—may prevent future behavior problems among high-risk children, yielding psychosocial benefits that endure through at least middle childhood without the need for additional intervention.